To visit our English website, Please click HERE

צוות המעבדה -

ד"ר ויקאס דוויבדי, פוסט-דוקטורנט

סלע יחזקאל, אגרונום

שרה שרף, מתנדבת
חברי המעבדה בעבר:
מעוז להב - תלמיד לתואר מוסמך, סיים בשנת 2004
שלי זפריאר - תלמיד לתואר מוסמך, סיים 2005
ד"ר ברוך צימרמן - פוסט דוקטורנט, סיים בשנת 2006
ליאור גולומב - תלמידה לתואר מוסמך, סיים בשנת 2007
מיכל הלפרט - תלמידה לתואר מוסמך, סיימה בשנת 2008
ד"ר דרור אבישר - פוסט דוקטורנט, סיים בשנת 2010
דוד שוורדשרף - סיים בשנת 2010
ד"ר יוני מסקוביץ' - פוסט דוקטורנט, סיים בשנת 2011
ד"ר אוקסנה רוגובוי - פוסט דוקטורנטית, סיימה בשנת 2012
אסנת אלטשולר - תלמידה לתואר מוסמך, סיימה בשנת 2013
אביב לוי - תלמיד לתואר מוסמך, סיים בשנת 2013
ראג' קומר תאפה - תלמיד לתואר מוסמך, סיים בשנת 2013
אינה מרדכייב - תלמידה לתואר בוגר, סיימה בשנת 2013
מרינה גרומברג - תלמידה לתואר מוסמך, סיימה בשנת 2014
סער לוינקרון - תלמיד לתואר בוגר, סיים בשנת 2014
אורי סררו - תלמיד לתואר מוסמך, סיים בשנת 2018
אבי אליהו - תלמיד לתואר מוסמך, סיים בשנת 2019
ספיר חגי - תלמידה לתואר בוגר, סיימה בשנת 2019
צביקה דומן - תלמיד לתאר שני, סיים בשנת 2020
מוחמד אבו-עביד- מהנדס מחקר, סיים בשנת 2020
גל הדס-ברנדווין- תלמידה לתאר שני, סיימה בשנת 2022
פן צאלה- תלמיד לתאר שני, סיים בשנת 2022
המחקר במעבדה מתמקד בנושאים הבאים:
ביולוגיה של התא ושלד התא
1. מיוזיונים מהצמח
מיוזינים הם חלבונים מוטוריים. בצמח חלבונים אלו משתייכים לשתי קבוצות עיקריות של מיוזינים לא קונבנציונאלים: מיוזין XI ומיוזין VIII, שניהם קרובים מבחינה פילוגנטית למיוזין V מיונקים. משפחת המיוזין בצמח המודל Arabidopsis thaliana כוללת 17 חברים: 13 מקבוצת מיוזין XI (מיוזין XI-A-K, XI-1 ו-XI-2) ו-4 מקבוצת מיוזין VIII (ATM1, ATM2, מיוזין VIII-A ו-VIII-B). ידוע שבני משפחת מיוזין XI מניעים את התנועתיות של אברונים גדולים, שלפוחיות קטנות, הרשת האנדופלזמית (ER) והציטופלזמה כולה בתאי הצמח. עם זאת, המנגנון העומד בבסיס התהליך אינו מובן במלואו. הסיבה לחוסר הבהירות היא שבעוד שהתנועתיות של גופיפי Golgi, פרוקסיזומים ומיטוכונדריה מופחתת בצמחים מוטנטיים של מיוזין xik, מיוזין XIK רקומביננטי מתפקד באורך מלא, מאוחה עם YFP, לא נצפה קשור לאברונים אלה. שלושה מנגנונים הוצעו כדי להסביר את אי הבהירות הזו: 1. מיוזין XI קושר שלפוחיות הובלה ספציפיות הנעות לאורך סיבי אקטין ויוצרות זרימה הידרודינמית של הציטופלזמה. כל שאר האברונים והשלפוחיות נעים על ידי זרימה זו ללא אינטראקציה ישירה עם מיוזין XI. 2. מיוזין XI נקשר ישירות לשלפוחיות והאברונים אך באופן חולף מאוד ומניע אותם רק למרחקים קצרים לאורך מסלולי אקטין, ולכן קשה מאוד ללכוד את הסיטואציה בעין המיקרוסקופ. 3. כמות מולקולות המיוזין הנקשרות בו זמנית לאברון נמוכה ומתחת רמה הניתנת לזיהוי במיקרוסקופ.
אנו חוקרים את האפשרות ששינויים מרחביים וזמניים לאחר תרגום של זנב מיוזין XI מסדירים את הקישור שלו לאברונים שונים. בנוסף, מיוזין פונקציונלי XIK-YFP רקומביננטי שבוטא במוטנט המשולש xik/xi2/xi1 נמצא בסיבי הקישור ובאזור האמצע של הפרגמופלסט בתאים מתחלקים הן במריסטמות של השורש והן במריסטמות של הנצר, דבר המצביע על תפקוד נוסף של מיוזין בחלוקת תאים. גם נושא זה הוא בתוכנית המחקר.
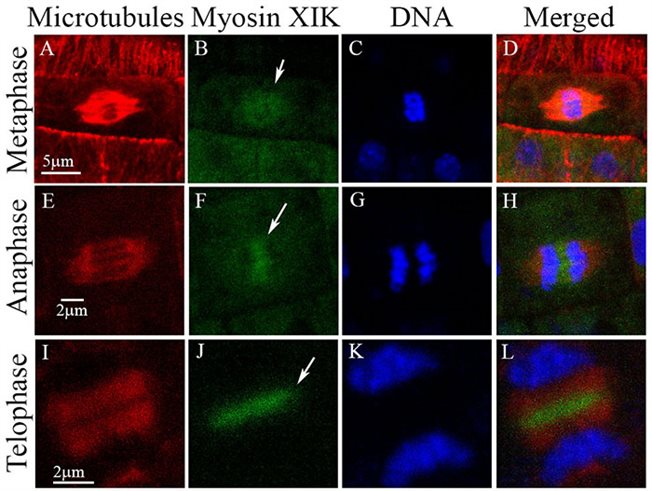
מיוזין XIK במנגנון חלוקת התא (Abu-Abied et al 2018)
שלד המיקרוטובולי בצמחים
שלד המיקרו-צינוריות הצמחיות (MTs) מאורגן לשני מבנים עיקריים: במהלך חלוקת התא, הוא יוצר את מנגנון חלוקת התא, אשר משתנה ברצף מטבעת הקדם-פרופאזה, אל סיבי הקישור ולאחר מכן הפרגמופלסט; במהלך שלב האינטרפאזה יוצרים סיבי המיקרוטובולי מערכים מקבילים מתחת לממברנת התא, המכונים מיקרוטובולי קורטיקליים. מיקרוטובולי קורטיקליים נשארים קשורים לממברנת התא לכל אורכם במהלך שלב האינטרפאזה. תחת המיקרוסקופ, מיקרוטובולי שאינם מסומנים נראים לעתים כחריצים שחורים בדפוס הפיזור של חלבונים מסומנים הקשורים לממברנה. על פי הסבר אחד, זה קורה מכיוון שמיקרוטובולי יכולים להגביל דיפוזיה רוחבית של חלבוני הממברנה על ידי יצירת כעין "גדרות" חוסמות. אנחנו הראינו לראשונה שמיקרוטובולי יכולים גם להתנגש בחלבונים הקשורים לממברנה ולשנות את הכיוון שלהם בהתאם, כך ששני המרכיבים משפיעים זה על הארגון של זה.
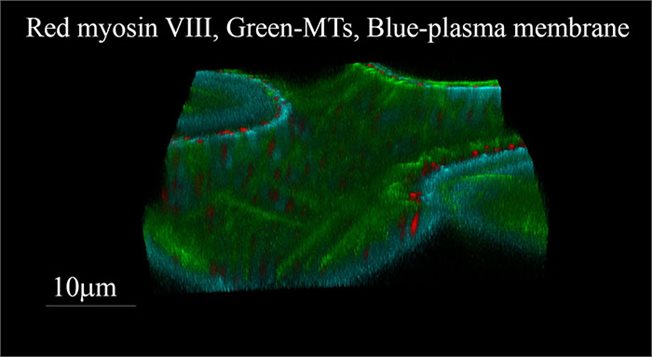
תצוגה תלת מימדית של תא אפידרמיס של Nicotiana benthamiana המבטא סמן למיקרוטובולי (GFP) ולזנב של מיוזין VIII (RFP).
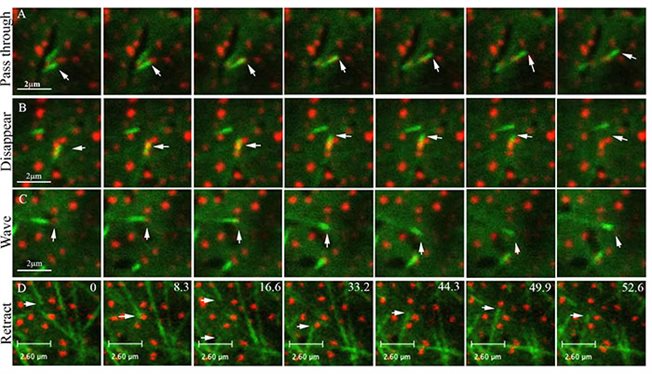
תרחישים שונים של אינטראקציה בין קצוות מתארכים של מיקרוטובולי מסומנים ב- EB1-YFP לבין זנבו של החלבון הממברנלי מיוזין VIII המסומן באדום (Bar-Sinai et al. 2022)
המנגנונים העומדים בבסיס התמיינות שורשים אדוונטיבים
שורשים אדוונטיבים נוצרים מרקמות שאינן שורש. השראת שורשים מיחורי גזע היא אחת הדרכים הנפוצות לריבוי וגטטיבי ומהווה שיטת עבודה חשובה בחקלאות. השרשת ייחורים חשובה מבחינה כלכלית בייעור, בתעשיית צמחי הנוי ובפיתוח כנות עילית להגדלת היבול, עמידות בפני מזיקים, מחלות ותנאי סביבה, בצמחים שונים, בעיקר עצי פרי אבל לא רק. למרות החשיבות הכלכלית והחקלאית הרבה של ריבוי וגטטיבי, עדיין קיים מחסום רציני בהשרשת ייחורים של צמחים יקרי ערך בעלי כושר השתרשות נמוך באופן טבעי או בכאלו שמאבדים יכולת זו במהלך ההתבגרות. הוכח שהתמיינות שורשים אדוונטיבים מבוקרת על ידי רשת מורכבת של העברת אותות ממגוון הורמונים צמחיים, שבה איתות מאוקסין ממלא תפקיד מרכזי בכל שלב בתהליך. מתן אוקסין לבסיס הייחור הוא הטיפול הנפוץ כדי לגרום להיווצרות שורשים אדוונטיבים, עם זאת, צמחים קשי השתרשות רבים אינם מייצרים שורשים בתגובה לטיפול כזה.
עבודתנו בנושא זה מתמקדת בשני תחומים: 1. פיתוח משפרי השתרשות חדשים, בשיתוף עם ד"ר רועי וינשטיין מאוניברסיטת תל-אביב. התרכובות החדשות שלנו נבדקות על יכולתן לשפר היווצרות שורשים אדוונטיבים בצמחים קשי השתרשות כגון עצי אקליפטוס, ארגן, כנות פקאן, גויאבה, מיני פרוטיאה ועוד. 2. התחום הנוסף כולל את הביולוגיה של התא העומדת בבסיס היווצרות שורשים אדוונטיבים. בתחום זה אנו עוסקים בשאלות הקשורות לתרומתם של שינויים בדופן התא להתמיינות של השורשים, כמו גם היבטים שונים של הובלת אברונים ממברנאלים בתא, אינטראקציות חלבון-חלבון ושינויים שלאחר תרגום בחלבונים שוינם, המשפיעים על היווצרות שורשים אדוונטיבים, או לחלופין על היווצרות קאלוס בבסיס הייחור.
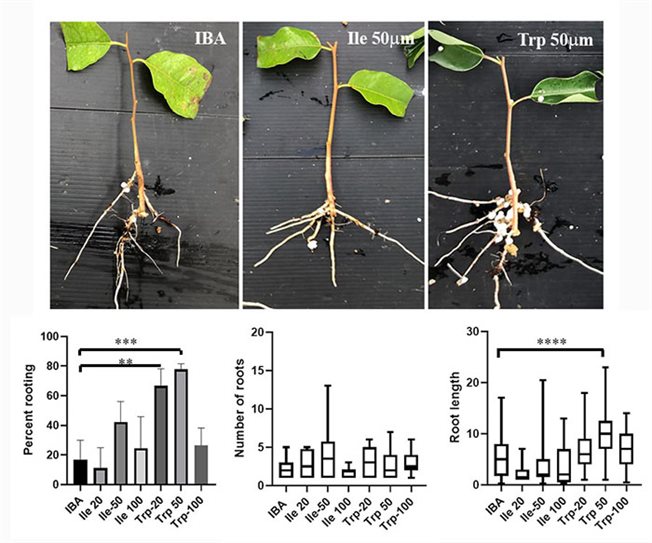
השתרשות ייחורים מעצי אקליפטוס גרנדיס בוגרים באמצעות תצמידים כמשפרי השתרשות חדשים

תאים ממויינים בקאלוס שנוצר בבסיס ייחור של כנת האבוקדו.
A. תאי עצה מעגליים.
B. רקמת שעם
C. רקמת אגירה (אור עובר).
D. רקמת אגירה (אור מקוטב).
E. ייחור כנת אבוקדו עם קאלוס ושורש בבסיס.

לרשימת הפרסומים עבור לאתר באנגלית